Navigation auf uzh.ch
Navigation auf uzh.ch
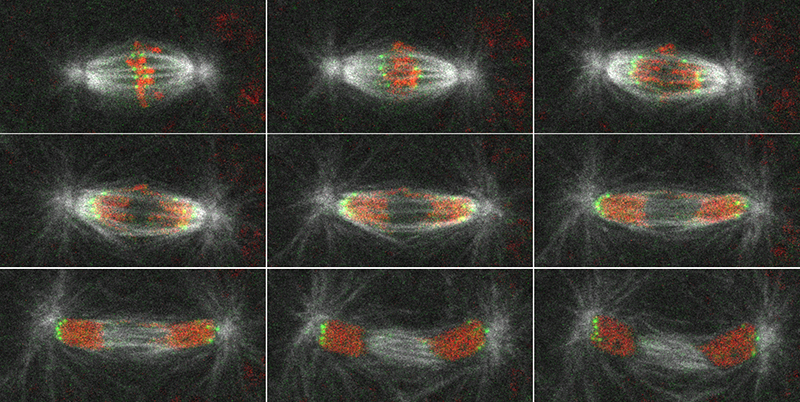
3.1 Mitotic cyclins and their functional specificity (Cyclin A, B und B3, Cdk1)
Cyclins are regulatory proteins that associate with cyclin-dependent protein kinases (cdk), permitting their activation. Mitotic cyclins bind to Cdk1. The activation of this kinase triggers entry into mitosis (chromosome condensation, nuclear envelope break down, mitotic spindle assembly). Conversely, exit from mitosis (mitotic spindle disassembly, nuclear envelope formation, chromosome decondensation) is dependent on inactivation of Cdk1. Inactivation results from the proteolytic degradation of the regulatory cyclin subunits. In Drosophila, we have identified three mitotic cyclin genes (Cyclin A, B and B3). We have isolated mutations in these conserved genes and have studied the resulting mitotic defects to define their specific functions.
3.2 Mitotic cyclin destruction (fizzy/CDC20, fizzy-related/CDH1, Mps1)
The rapid destruction of mitotic cyclins by the ubiquitin-dependent protein degradation system is essential for progression through mitosis. This mitotic cyclin destruction is only initiated when all chromosomes are properly aligned in the mitotic spindle (spindle assembly checkpoint, SAC). After mitotic cyclin destruction, exit from mitosis (mitotic spindle disassembly, nuclear envelope formation, chromosome decondensation) can proceed. Destruction of mitotic cyclins continues throughout G1 phase and is only inactivated after entry into S phase. We have identified two conserved genes in Drosophila, fizzy and fizzy-related, which play a crucial role in regulating cyclin destruction. We have studied the role of the Mps1 kinase in the spindle assembly checkpoint (SAC).
3.3 Centromeres, kinetochores and sister chromatid separation during mitosis (centromere kinetochore components: Cenp-A/Cid, Cenp-C, KMN network; Separase, cohesin, C(2)M)
Cohesion of sister chromatids in mitotic chromosomes is essential for the correct bipolar orientation of sister kinetochores in the bipolar spindle during mitotic prophase. However, sister chromatids have to be separated at the metaphase-anaphase transition to allow their segregation to the poles. We have identified two Drosophila genes, three rows and pimples, which are specifically required for sister chromatid separation during mitosis. The gene products (THR, PIM) associate with the endoprotease Separase (SSE), which permits sister chromatid segregation by cleaving a subunit of the cohesin complex. The Separase complex interacts genetically with centromere proteins which motivated our characterization of centromere and kinetochore proteins.
3.4 G1 cyclins and the exit from mitotic proliferation (Cyclin D/Cdk4/6, Cyclin E/Cdk2, dacapo)
G1 cyclins (cyclin D and E) in association with distinct cdks (Cdk4/6, Cdk2) control entry into the cell division cycle. Conversely, the arrest of cell proliferation at the correct developmental stage depends on timely inactivation of these Cyclin-Cdk complexes. With a genetic screen, we have identified the Drosophila dacapo gene which encodes a CIP/KIP-type inhibitor of Cyclin E-Cdk2 complexes. Dacapo expression is strongly upregulated during the final division cycle and thereby enforces a timely cell proliferation arrest. By generating mutant alleles and analysis of phenotypic consequences, we have also characterized the role of Cyclin D-Cdk4/6 complexes in the control of cell proliferation during Drosophila development.
Key publications: