What is Caenorhabditis elegans?
The roundworm (Nematode) Caenorhabditis elegans has become a popular model organism in Biology since it was introduced by Sydney Brenner in the early 1970ies. The broad range of genetic and molecular techniques that are available in Caenorhabditis elegans is one of the main reasons why this model organism has become so popular among many biologists. Today, over 3000 scientists worldwide are investigating the biology of Caenorhabditis elegans.
The research on this approximately 1 mm long roundworm that consists of fewer than 1000 body cells has revealed some fundamental and ubiquitous biological processes. One example is the phenomenon of programmed cell death (Apoptosis). It was first described in Caenorhabditis elegans and found to be a genetically controlled process. Today, we know that programmed cell death occurs in most if not all animals and plants. Another example is the inactivation of gene expression through double-stranded RNA (RNAi or RNA interference) that was discovered in Caenorhabditis elegans. In addition, the study of Caenorhabditis elegans has provided important insights into the origins of human diseases such as Cancer or Alzheimer’s disease. Several human genes that play a key role in the formation of these diseases have their counterparts (homologs) in Caenorhabditis elegans. By studying the Caenorhabditis elegans homologs of human disease genes, we can investigate the molecular mechanisms that cause the corresponding diseases in humans. Such experiments may ultimately lead to the development of new therapies and drugs.
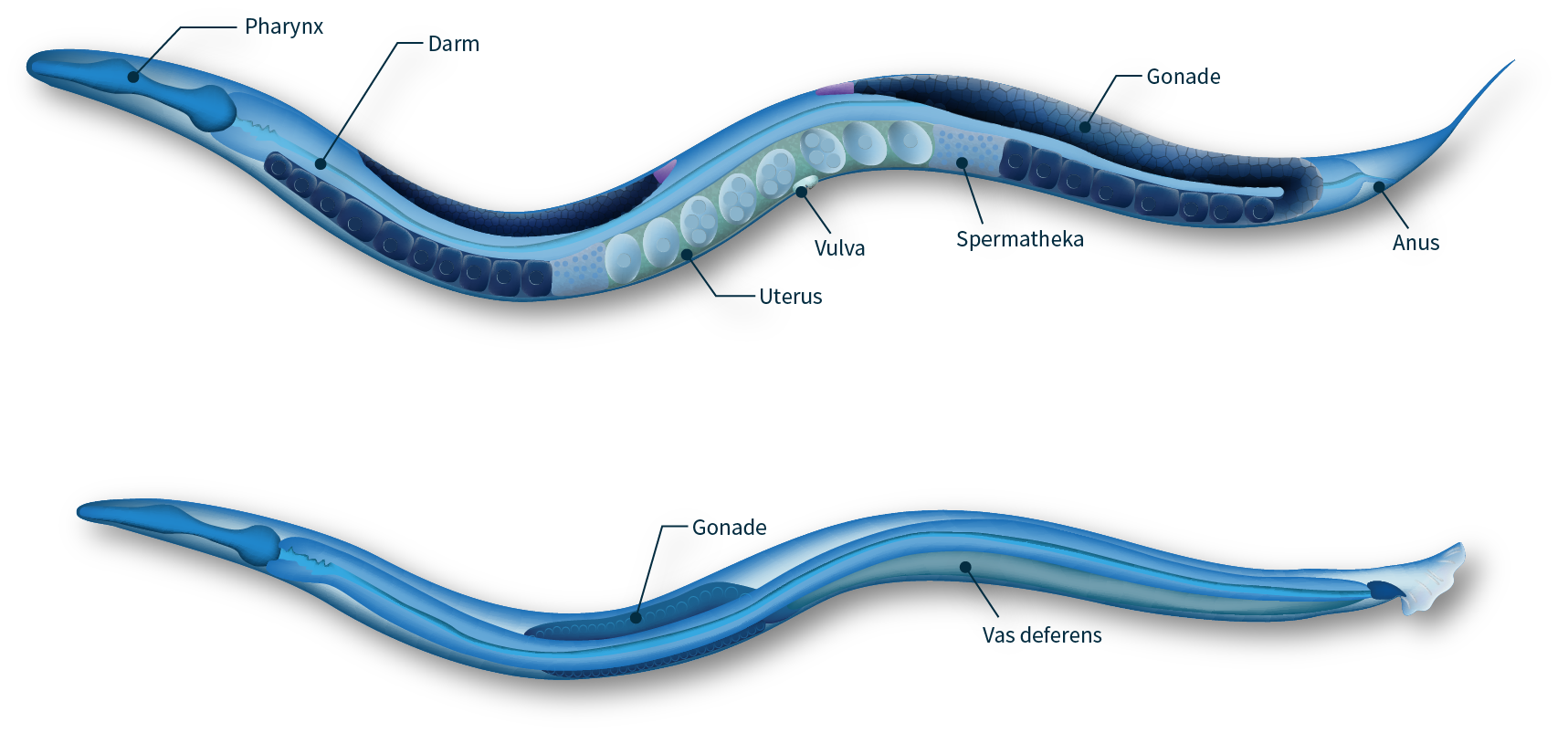
- worm (PNG, 399 KB)
- Anatomy of adult hermaphrodite and male C. elegans
{kind=link}
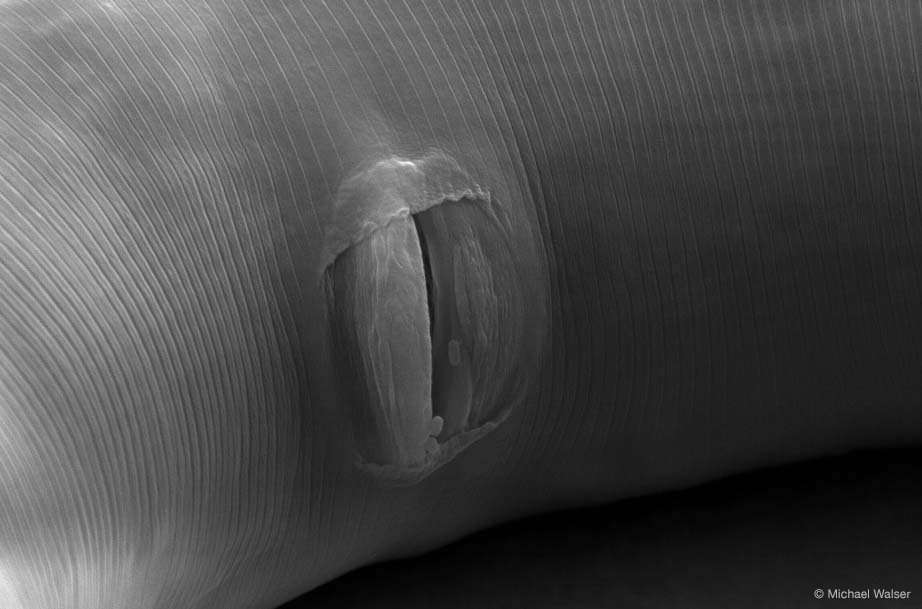
- vulva (JPG, 109 KB)
- Scanning electron microscopy image of an adult C. elegans vulva
{kind=link}
Vulval development
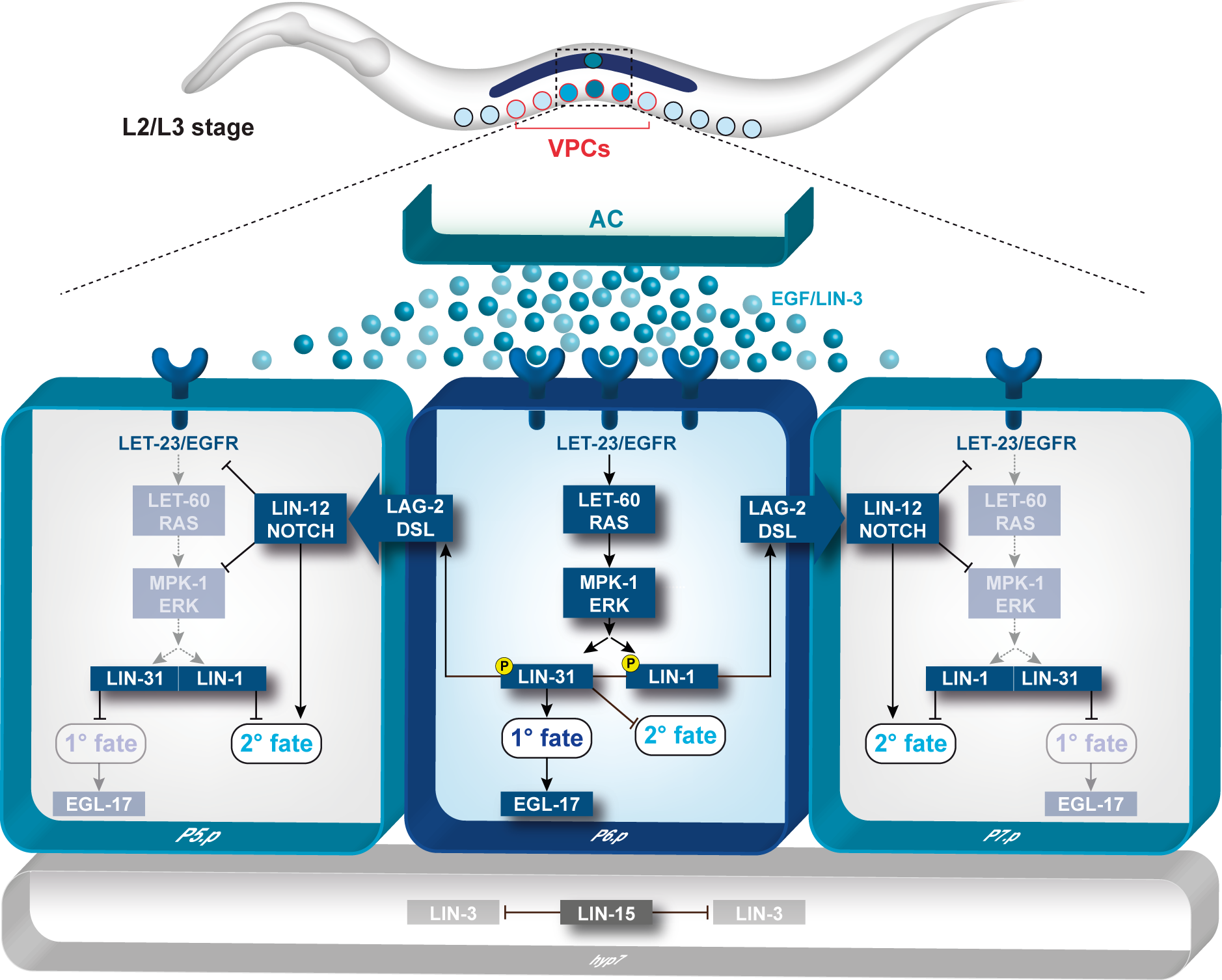
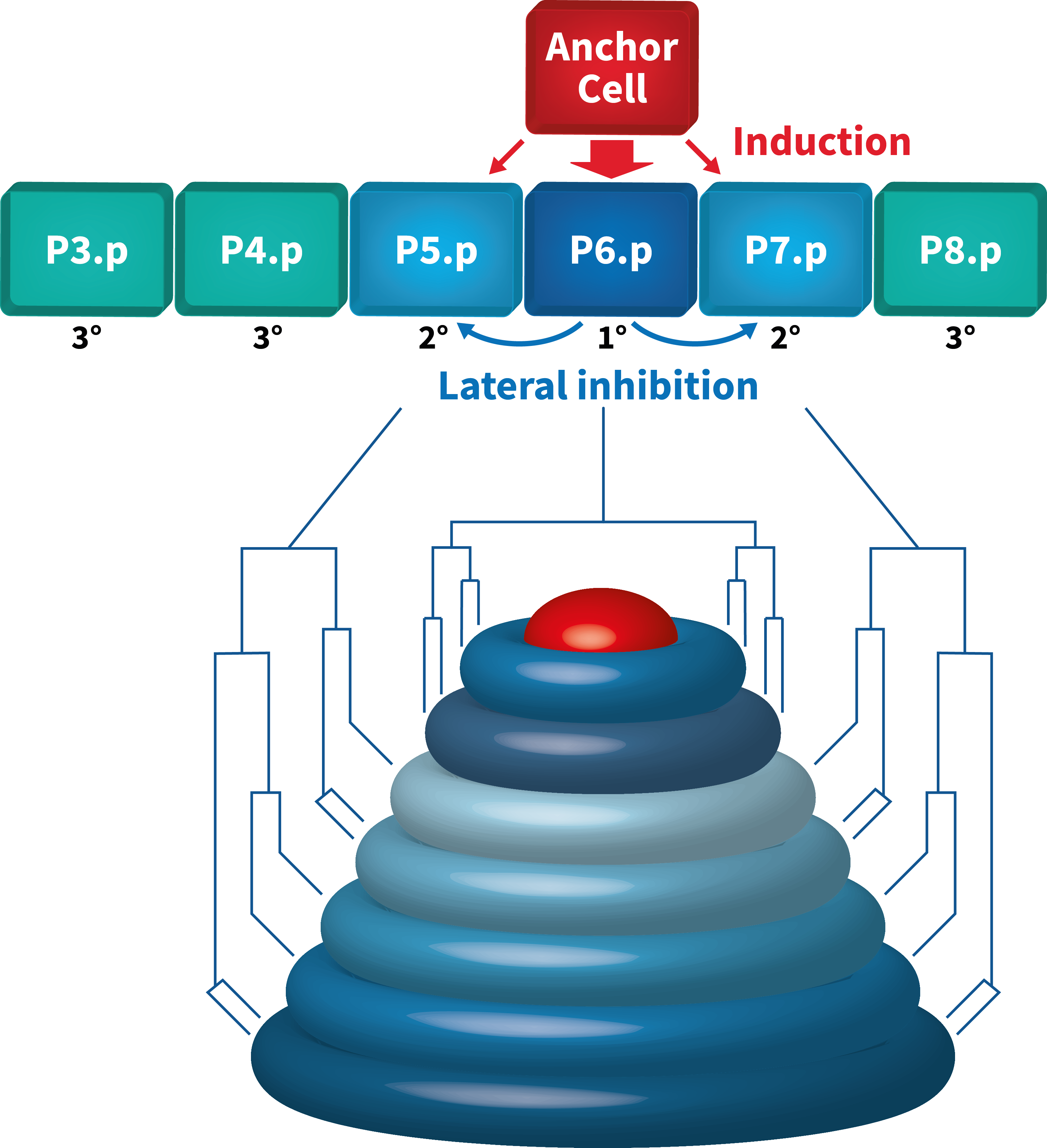
The hermaphrodite vulva is a simple organ consisting of 22 cells that are formed during the postembryonic (larval) development. Towards the end of the first larval stage, a Wingless (Wnt) signal specifies the identity of the six equivalent vulval precursor cells (VPCs) P3.p through P8.p, that form the vulval equivalence group.
The Wnt signal induces the expression of the Hox gene lin-39 that specifies the vulval equivalence group and maintains the VPCs as polarized epithelial cells. Thus, the VPCs are competent to respond to an inductive signal that is produced by the gonadal anchor cell (AC) during the second and third larval stages. By the third larval stage, the fates of the VPCs have been specified by the combined action of the EGFR/RAS/MAP kinase and the DELTA/NOTCH signaling pathways. The AC produces the LIN-3 epidermal growth factor (EGF) that activates the receptor tyrosine kinase LET-23 (the EGF receptor) in the closest VPC P6.p. This inductive signal is transduced in P6.p by the conserved EGFR/RAS/MAP kinase signal transduction pathway where it specifies the primary (1°) vulval fate. In response to the inductive AC signal, P6.p generates a lateral signal that activates the NOTCH signaling pathway in its neighbors P5.p and P7.p. NOTCH signaling first inhibits the induction of the 1° fate in a process termed lateral inhibition. In a second step, the NOTCH pathway specifies the secondary (2°) fate in P5.p and P7.p. The VPCs that receive neither inductive nor lateral signals adopt the tertiary (3°) fate. They fuse with the epidermis (hyp7) after undergoing one round of cell divisions. After the 1° and 2° fates have been specified, the VPCs execute a stereotypic pattern of cell divisions, and their descendants undergo extensive morphogenesis during the third and fourth larval stages. The cells migrate and invaginate to form the ventral opening. Several cell fusions between the differentiated vulval cells occur to form a tubular organ that connects to the uterus to allow egg-laying.
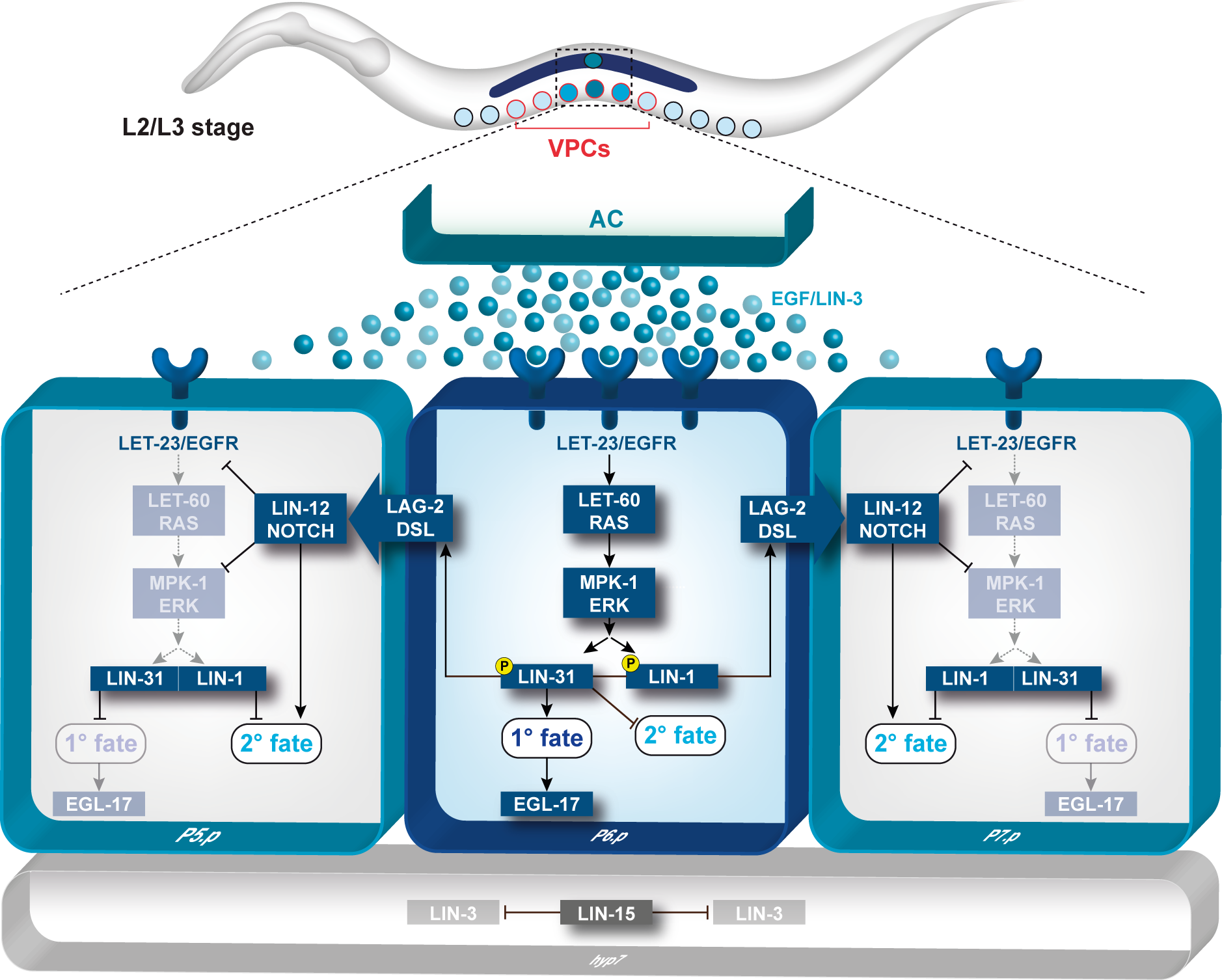
- toroids (PNG, 522 KB)
- Signal transduction during C. elegans vulval development
{kind=link}
- toroids (PNG, 736 KB)
- Toroid formation
{kind=link}
Human disease genes: The wrong signal at the wrong time
Several human genes that are involved in the formation of various diseases have been identified. In the affected patients, these genes often carry mutations that perturb the normal function of the proteins they encode. It was soon realized that many of the so-called “human disease genes” have homologs in most animals, where these genes perform essential functions during normal development.
Well-studied examples are the oncogenes Ras, which is mutated in a large portion of epithelial cancers(carcinoma), or the ErbB-2 gene, which is activated through mutations in breast cancer. On the other hand, the tumor suppressor genes Rb (in retinoblastoma), Patched (in basal cell carcinoma, a form of skin cancer) or APC (in colorectal, intestinal cancer) are often inactivated in those tumors. During normal development, the same genes function in important inter-cellular signaling pathways: The ErbB-2 gene, for example, encodes a receptor protein that becomes activated when it binds to specific growth factors at the cell surface. The RAS protein is localized at the inner side of the plasma membrane, where it acts as a regulatory switch: RAS is activated by the signals from growth factor receptors at the cell surface and transmits the growth-inducing signals into the cytoplasm of the cells. The mutations, which are frequently found in the human RAS oncogenes, lock this switch in the “ON” position.
Key publications:
Signal transduction during C. elegans vulval development: a NeverEnding story.
Schmid T, Hajnal A.
Curr Opin Genet Dev. 2015 Jun;32:1-9. doi: 10.1016/j.gde.2015.01.006. Epub 2015 Feb 9. Review.